1. INTRODUÇÃO
O sistema genital masculino é composto pelas gônadas (testículos) e órgãos genitais acessórios,, incluindo os ductos excretores genitais e as glândulas acessórias (glândulas seminais - bilaterais -, próstata - única e mediana - e glândulas bulbouretrais - também bilaterais), além da genitália externa constituída pelo pênis e pelo escroto.
Os testículos, por sua vez, possuem duas funções principais: a espermatogênese e a esteroidogênese, ou seja, produção de espermatozoides e síntese de andrógenos, respectivamente. O principal andrógeno produzido pelos testículos é a testosterona e, juntamente com o seu metabólito, a di-hidrotestosterona, são de fundamental importância para a fisiologia do corpo masculino desde o período embrionário até a vida adulta, visto que eles exercem papel central no dimorfismo sexual e caracteres sexuais masculinos,, além de exercer papel sobre o processo de espermatogênese.
As glândulas acessórias fabricam secreções e lançam nos ductos genitais, os quais quando impulsionados por contrações musculares (ejaculação), movem o sêmen em direção ao exterior através do pênis. Além de auxiliar no deslocamento dos espermatozoides, essas secreções ofertam nutrientes aos espermatozoides enquanto estes se encontram no aparelho reprodutor masculino.
Por fim, o sêmen é composto pelos espermatozoides e pelas secreções provenientes das glândulas acessórias. O líquido seminal contém nutrientes para os espermatozoides e fornece proteção para estes durante sua passagem pelo sistema de ductos excretores e trato genital feminino. Possui pH alcalino, em torno de 7,7, o que ajuda a neutralizar o pH ácido da vagina e da uretra do homem. Os principais componentes do sêmen advêm das secreções das glândulas seminais (65 a 75%) e da próstata (25 a 30%).
2. TESTÍCULOS
Os testículos são as gônadas masculinas, que integram tanto o sistema genital quanto o endócrino. Eles se localizam fora da cavidade abdominal, no interior do saco escrotal (ou escroto). Os testículos e os funículos espermáticos associados são revestidos por camadas teciduais distintas adquiridas durante a descida ou migração das gônadas masculinas a partir de sua posição retroperitoneal original na cavidade abdominal para o saco escrotal. Durante a vida intrauterina, no início da migração testicular, começa o desenvolvimento do escroto e do processo vaginal, uma projeção do peritônio que formará a túnica vaginal, sendo constituída em uma camada parietal exterior e uma camada visceral interna, sendo responsável pela cobertura da túnica albugínea nas suas porções laterais e anterior do testículo e epidídimo.
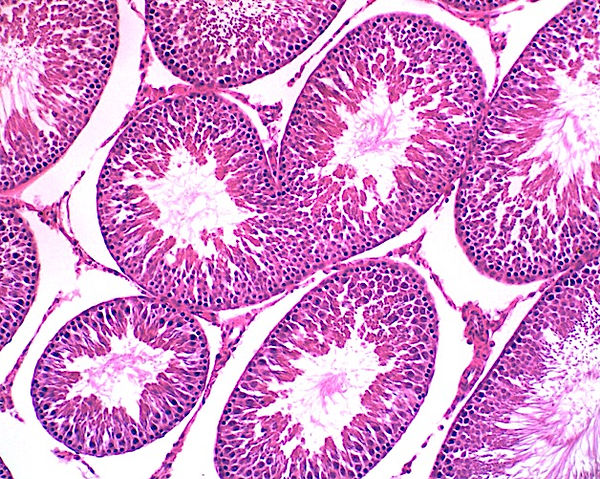
Cada testículo está envolto por uma camada espessa de tecido conjuntivo denso modelado, a túnica albugínea. Ela é mais densa na superfície posterior dos testículos para formar o mediastino do testículo, do qual se originam os delgados septos fibrosos. Os septos ao penetrarem no testículo, divide-o em compartimentos piramidais chamados lóbulos testiculares. Cada testículo é dividido em aproximadamente 250 lóbulos que se originam a partir dessa cápsula de tecido conjuntivo. Esses septos são incompletos e muitas vezes há comunicação entre os lóbulos. Cada lóbulo do testículo é composto de um a quatro túbulos seminíferos, local onde os espermatozoides são produzidos. Os túbulos seminíferos são envolvidos por um tecido conjuntivo frouxo rico em vasos linfáticos e sanguíneos, nervos e células de Leydig (intersticiais) que são as responsáveis pela produção de testosterona. Além disso, o tecido intersticial do testículo é indispensável para a nutrição das células dos túbulos seminíferos, a produção de andrógenos e o transporte destes hormônios para a circulação sistêmica.
Devido ao seu comprimento considerável, cada túbulo no lóbulo forma uma alça e é altamente contorcido, dobrando-se sobre si mesmo no lóbulo. As extremidades dos lóbulos se encontram próximo ao mediastino do testículo, onde assumem um novo trajeto reto e curvo. Este segmento do túbulo seminífero é denominado túbulo reto.
Os túbulos retos fazem a conexão entre os túbulos seminíferos com uma rede de canais anastomosados revestido por um epitélio simples pavimentoso ou cúbico, constituindo a rede testicular no mediastino do testículo. De 10 a 20 ductos eferentes conectam à rede testicular a porção proximal do ducto do epidídimo. Os ductos eferentes são constituídos de células epiteliais cuboides que se alternam com células ciliadas que batem seus cílios na direção do epidídimo. Assim, o batimento dessas células cria um fluxo que transporta os espermatozoides em direção ao epidídimo. Os ductos eferentes vão se fundindo para formar o ducto do epidídimo.
Corte sagital de testículo e epidídimo
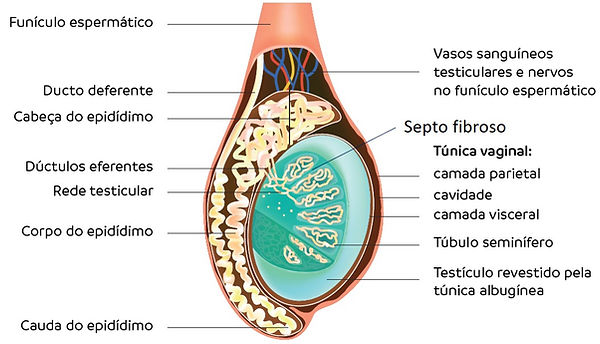
Imagem adaptada de BORGES, G.R., 2019.

2.1 TÚBULOS SEMINÍFEROS
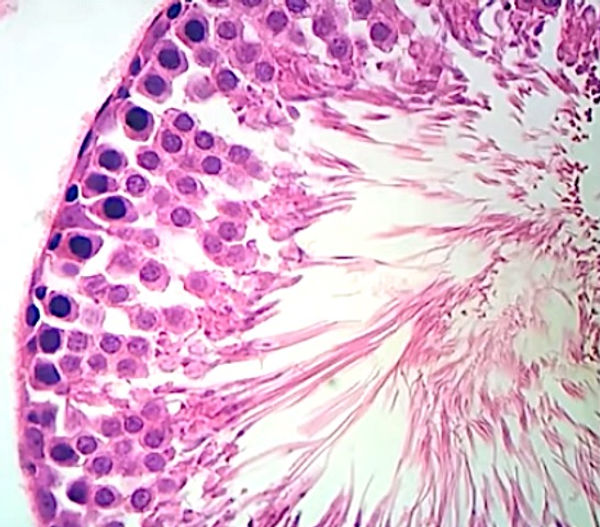
A parede dos túbulos seminíferos é formada por um complexo epitélio estratificado, apoiado sobre uma lâmina basal. O túbulo é envolvido por uma bainha de tecido conjuntivo frouxo, rica em células mioides (células contráteis peritubulares) e fibras colágenas. As contrações rítmicas das células mioides criam ondas peristálticas que impulsionam os espermatozoides e o líquido testicular através dos túbulos seminíferos até o sistema de ductos excretores. O epitélio é constituído por duas populações celulares: as células da linhagem seminífera, ou espermatogênica, e as células de Sertoli ou de sustentação, não proliferativas. Em um túbulo seminífero, as células da linhagem germinativa se encontram em vários estágios da espermatogênese.
-
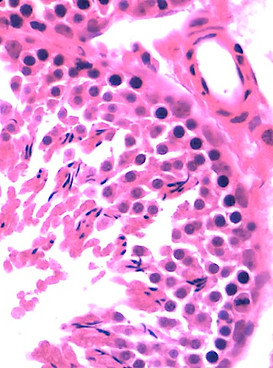
Células espermatogênicas (epitélio germinativo): são as células que se replicam e se diferenciam em espermatozoides maduros. As células da linhagem germinativa se encontram em vários estágios da espermatogênese. As células mais próximas à lâmina basal com núcleos esféricos são as espermatogônias. As células maiores com núcleos esféricos, mas com uma cromatina nuclear característica, contendo grumos alongados grosseiros, correspondem aos espermatócitos primários. Os espermatócitos secundários, já haploides, são raramente vistos; tão logo se formam, eles já se dividem e dão origem às espermátides. As espermátides, que são caracterizadas como iniciais ou tardias, não se dividem, mas apenas sofrem maturação em espermatozoides, os quais são liberados no lúmen dos túbulos seminíferos e carreados para diante nas vias espermáticas.
-
Células de Sertoli: desempenham um papel fundamental no suporte e na maturação dos espermatozoides. Após a puberdade, elas constituem cerca de 10% das células no epitélio seminífero. Essas células cilíndricas, com limites que são difíceis de distinguir, se estendem da lâmina basal até o lúmen do túbulo seminífero. As células de Sertoli não se replicam após o período da puberdade e são reconhecidas na microscopia principalmente pelos seus núcleos situados na base dos túbulos seminíferos. Os núcleos têm formato vesicular, claros (com eucromatina), triangulares e frequentemente contêm um nucléolo evidente. O citoplasma dessas células não é visto com muita nitidez, logo, os limites dessas células são mal definidos. Células adjacentes são unidas por junções de oclusão no perímetro basal, de modo que a parede dos túbulos seminíferos seja dividida em um compartimento basal e um compartimento adluminal. As junções de oclusão entre as células de Sertoli constituem a barreira hematotesticular, a qual promove a separação entre as espermatogônias e os espermatócitos primários no compartimento basal e os espermatócitos secundários e espermátides no compartimento adluminal. O conteúdo dos túbulos seminíferos fica, portanto, isolado de antígenos circulantes, consequentemente protegendo espermatócitos e espermátides de reações autoimunológicas e de substâncias advindas com o sangue. As células de Sertoli fagocitam resquícios de espermátides e secretam o fluido testicular e outras substâncias, incluindo a proteína de ligação a andrógenos, essencial para a sobrevivência dos espermatozoides (ABP) que concentrará a testosterona no compartimento luminal do túbulo seminífero. Nesse compartimento, a testosterona em concentrações elevadas é vital para a maturação dos espermatozoides. A atividade secretora das células de Sertoli é regulada por mecanismos de “feedback” ou alças de retroalimentação. Na superfície dessas células há receptores de FSH (hormônio folículo estimulante) e de testosterona que exercem um papel regulador no processo de espermatogênese. Para “fechar” o mecanismo de retroalimentação, as células de Sertoli secretam uma gama de substâncias endócrinas, dentre elas a inibina, que inibe a liberação de FSH pela adeno-hipófise.
TESTÍCULO - H.E. - 40x
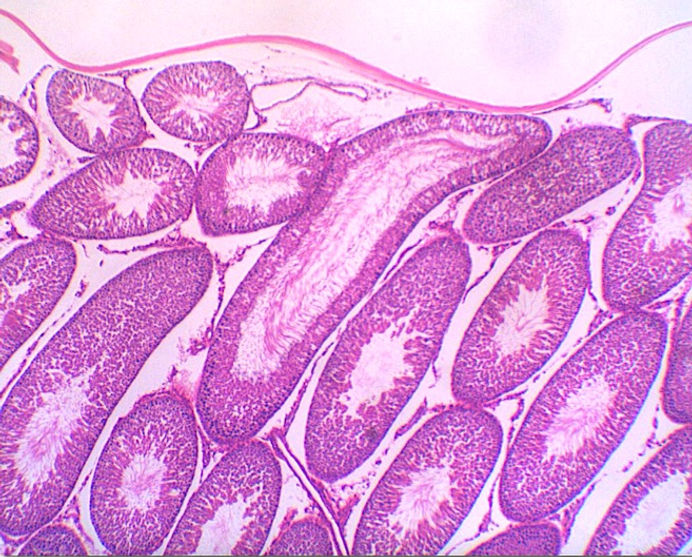
TÚBULO SEMINÍFERO - H.E. - 400x
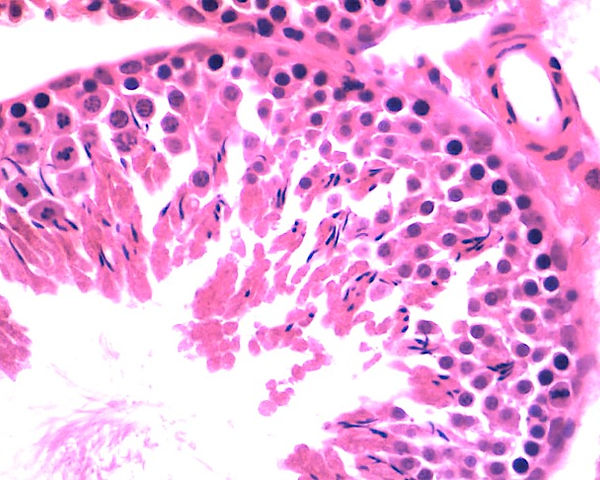
TÚBULO SEMINÍFERO - H.E. - 400x
2.2 TECIDO INTERSTICIAL
No tecido conjuntivo frouxo em meio aos túbulos seminíferos, estão presentes agregados de células de Leydig (intersticiais). Seu citoplasma ligeiramente acidófilo (eosinofílico) e vacuolizado se deve à remoção das gotículas lipídicas durante o processamento histológico, uma vez que elas armazenam colesterol para a síntese de testosterona.
A testosterona apresenta um papel importante a depender do estágio de desenvolvimento do indivíduo, sendo indispensável durante o desenvolvimento embrionário, a maturação sexual e a fase reprodutiva.
TESTÍCULO - H.E. - 100x
TESTÍCULO - H.E. - 400x
2.3 ESPERMATOGÊNESE
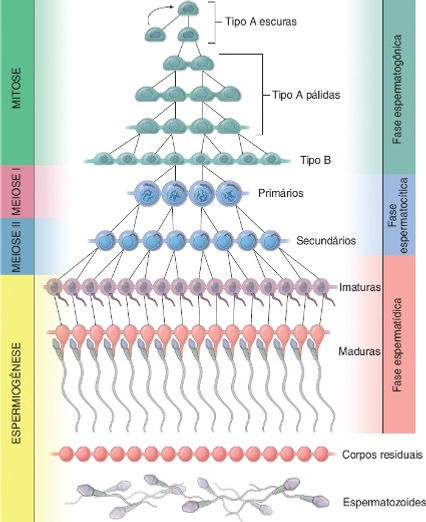
A espermatogênese envolve uma série complexa de eventos. Inicia-se pouco antes da puberdade, sob a influência dos níveis crescentes de gonadotropinas hipofisárias, e prossegue durante toda a vida. A espermatogênese é dividida em três fases distintas (espermatogônica, espermatocítica e espermatídica):
SUBSTITUIR (FELIPE)
• Fase espermatogônica: as espermatogônias dividem-se por mitose. A partir desse ponto, as células recém formadas podem seguir dois caminhos: manter-se como células-tronco de outras espermatogônias, recebendo a denominação de espermatogônias do tipo A, ou sofrerem um processo de diferenciação para se tornarem espermatogônias do tipo B. Ocorre nas espermatogônias de tipo B sucessivos ciclos mitóticos nos quais as células filhas não se separam por completo, dando origem aos espermatócitos primários. Estas células, por sua vez, se mantêm unidas por pontes citoplasmáticas até o fim do processo de espermatogênese.
• Fase espermatocítica (meiose): os espermatócitos primários sofrem duas divisões meióticas que reduzem tanto o número de cromossomos quanto a quantidade de DNA, e que também produzem células haploides, denominadas espermátides. Esses espermatócitos replicam o seu DNA logo após a sua formação e antes do início da meiose, de modo que cada espermatócito primário contém o número cromossômico normal (2n, 46 cromossomos). Como cada cromossomo é constituído por duas cromátides-irmãs, os espermatócitos primários contêm o dobro de DNA (4d, 4 cromátides). A meiose I (reducional) resulta em redução tanto do número de cromossomos (de 2n, 46 cromossomos para 1n, 23 cromossomos) quanto da quantidade de DNA para um número haploide (de 4d, 4 cromátides para 2d, 2 cromátides); por conseguinte, espermatócitos secundários têm um número haploide de cromossomos (1n, 23 cromossomos) e uma quantidade 2d (duas cromátides) de DNA. Como a meiose II (equacional) não é precedida por replicação do DNA, as espermátides, que são formadas após essa divisão, também terão um número haploide (1n, 23 cromossomos) de cromossomos, contendo, cada um deles, uma única cromátide (1d).
• Fase espermatídica (espermiogênese): as espermátides sofrem substancial remodelação celular, à medida que se diferenciam em espermatozoides maduros. Cada espermátide, que resulta da segunda divisão meiótica, apresenta um conteúdo de DNA haploide (1n, 23 cromossomos) e um número de cromossomos (1d, 1 cromátide) representado por 22 autossomos e por um cromossomo X ou Y. Não ocorre outra divisão. As espermátides haploides sofrem um processo de diferenciação que produz espermatozoides maduros, que também são haploides. A condição diploide (2d) normal é restaurada, quando um espermatozoide fertiliza um oócito.
A ESPERMIOGÊNESE é composta por três etapas com um certo grau de complexidade que resultam na produção de um espermatozoide maduro. Estas etapas são as seguintes:
1. Etapa do complexo de Golgi: as espermátides apresentam um complexo de Golgi bem desenvolvido. Nessa estrutura, acumulam-se os grânulos pró-acrossômicos que posteriormente irão se fundir para formar o grânulo acrossômico no interior da vesícula acrossômica. Ademais, nessa etapa, os centríolos migram para próximo da superfície celular em oposição a vesícula acrossômica, iniciando a construção do axonema, ou seja, o agrupamento de microtúbulos que formam o eixo central do flagelo do espermatozoide.
2. Etapa do acrossomo: a vesícula acrossômica se alonga sobre a metade anterior do núcleo, se assemelhando a um capuz, e por fim se tornando o acrossomo. Essa estrutura, contém diversas enzimas hidrolíticas, por exemplo, hialuronidase, neuraminidase e fosfatase ácida. Tais enzimas são de grande relevância para os espermatozoides, visto que elas são capazes de dissociar as células que compõem a corona radiata do ovócito. Paralelamente a esse processo, as mitocôndrias começam a se acumular na porção proximal do flagelo, formando a peça intermediária. A organização dessas organelas é estratégica, tendo em vista que as mitocôndrias atuam gerando energia e esta será utilizada para a movimentação do flagelo, já que o movimento flagelar resulta da interação entre os microtúbulos, os ATPs e a dineína (uma proteína com atividade de ATPase). Por fim, o núcleo das espermátides torna-se mais condensado e alongado, além de ficar voltado para a base do túbulo seminífero enquanto o flagelo se projeta em direção ao lúmen.
3. Etapa de maturação: nesse processo, há a formação dos corpos residuais, que nada mais são que partes perdidas do citoplasma das espermátides. Esses corpos residuais, posteriormente serão fagocitados pelas células de Sertoli e os espermatozoides maduros serão liberados no lúmen do tubo. A partir desse momento, eles serão transportados ao epidídimo, por meio do fluído testicular, no qual sob condições adequadas terminarão o processo de maturação. O fluido testicular é composto por esteroides, proteínas, íons, entre outros.
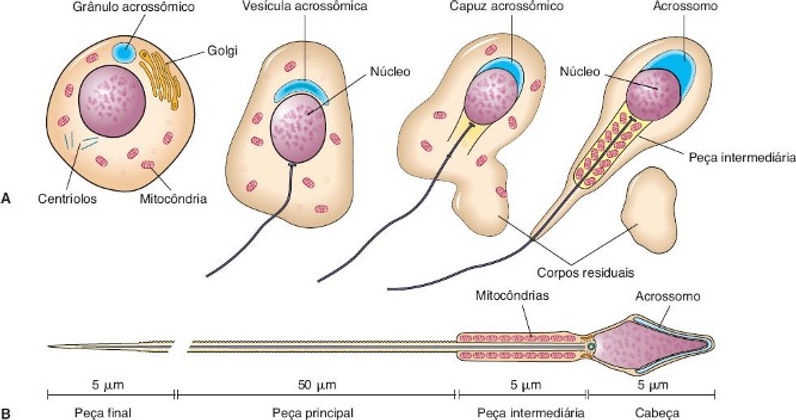
Imagem retirada de Junqueira e Carneiro, 2017.
Como descrito anteriormente, nos cortes histológicos do testículo, as células mais próximas à lâmina basal com núcleos esféricos são as espermatogônias. As células maiores com núcleos esféricos, mas com uma cromatina nuclear característica, contendo grumos alongados grosseiros, correspondem aos espermatócitos primários. A maioria dos espermatócitos são encontrados na fase de prófase, pois essa fase dura aproximadamente 22 dias. Ademais, os espermatócitos secundários não possuem fácil visualização, visto que estes permanecem por um curto período de tempo em interfase e após a sua formação, entram na segunda etapa de divisão da meiose, dando origem as espermátides.
As espermátides, que são caracterizadas como iniciais ou tardias, não se dividem, mas apenas sofrem maturação em espermatozoides, por sua vez, possuem como características que a distinguem das demais células: (1) um pequeno tamanho; (2) núcleos com cromatina condensada e de formato variável; (3) localizado próximo ao lúmen dos túbulos seminíferos.
Na conclusão da espermatogênese, as espermátides sofrem sua maturação final se diferenciando em espermatozoides que são liberadas no lúmen do túbulo seminífero, durante um processo denominado espermiação, pelas células de Sertoli ou de sustentação. Os espermatozoides são liberados no lúmen dos túbulos seminíferos e carreados para diante nas vias espermáticas.
2.4. REGULAÇÃO HORMONAL DA ESPERMATOGÊNESE
A função normal dos testículos depende de hormônios que atuam por meio de vias endócrinas e parácrinas. Os dois principais hormônios adeno-hipofisários que participam da espermatogênese são o Hormônio luteinizante ou LH e o hormônio folículo estimulante ou FSH.
A função endócrina do testículo reside principalmente na população de células de Leydig, que estimuladas pelo LH, sintetizam e secretam o principal andrógeno circulante, a testosterona. Quase toda a testosterona é produzida pelos testículos; menos de 5% desta é produzida pelas glândulas suprarrenais. Quando a testosterona deixa as células de Leydig, cai na corrente sanguínea e nos capilares linfáticos, e então cruza o tecido peritubular a fim de alcançar o epitélio seminífero.
O FSH age nas células de Sertoli, promovendo a síntese e a secreção de proteína ligante de andrógeno (ABP). Como a testosterona é produzida pelas células intersticiais (localizadas fora dos túbulos seminíferos) e levando em consideração que são necessárias concentrações elevadas de testosterona dentro dos túbulos, as proteínas ABP são de extrema necessidade, visto que permitem uma concentração elevada de testosterona dentro dos túbulos seminíferos, condição muito importante para estimular a espermatogênese.
As células de Sertoli secretam continuamente nos túbulos seminíferos um fluido que é transportado na direção dos ductos genitais e é usado para transporte de espermatozoides. A secreção de ABP pelas células de Sertoli é controlada pelo FSH e por testosterona. Células de Sertoli podem converter testosterona em estradiol e também secretam um peptídio chamado inibina, que suprime a síntese e a liberação de FSH na hipófise (retroalimentação negativa).

SUBSTITUIR FELIPE
3. EPIDÍDIMO
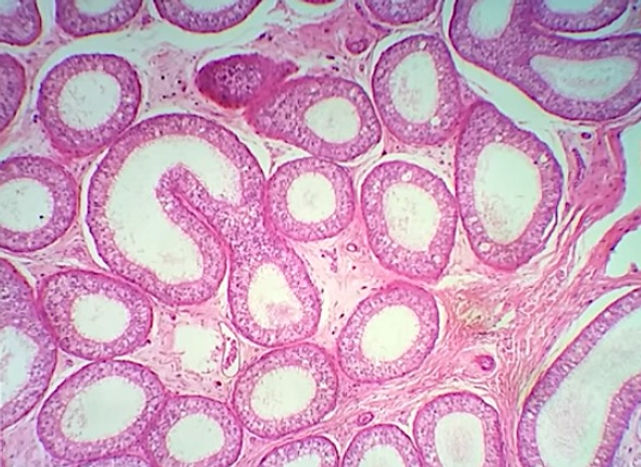
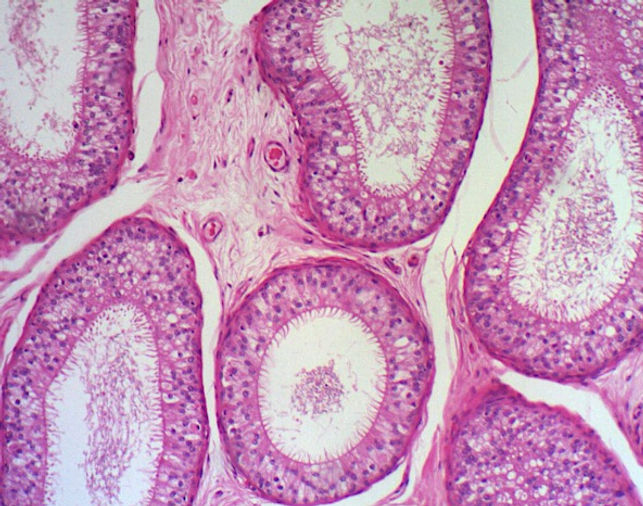
O epidídimo é uma estrutura em formato de meia-lua, localizada ao longo das superfícies superior e posterior do testículo. Consiste nos dúctulos eferentes e no ducto do epidídimo (ducto epididimário). Apresenta vasos, músculo liso e uma cobertura de tecido conjuntivo. O ducto do epidídimo é um tubo altamente espiralado. O epidídimo é dividido em cabeça, corpo e cauda. Os dúctulos eferentes que se ligam com o testículo para trazer os espermatozoides, ocupam a cabeça, enquanto o ducto do epidídimo ocupa o corpo e a cauda. O ducto do epidídimo é longo, tortuoso e enovelado. Devido à sua tortuosidade, os cortes histológicos podem mostrar segmentos do ducto em sentidos longitudinal, transversal ou oblíquo.
Dentro do epidídimo, os espermatozoides amadurecem, adquirindo motilidade e capacidade de fertilizar um oócito. Essa maturação é dependente de andrógenos e ocorre à medida que o espermatozoide recém-formado percorre o ducto do epidídimo (14 dias). Durante o processo de maturação do espermatozoide, a cabeça do espermatozoide é alterada de modo reversível pela adição do fator de descapacitação associado à superfície. Esse procedimento, chamado de descapacitação, impede a capacidade de fertilização do espermatozoide, mas de modo reversível. Quando o espermatozoide é liberado no trato genital da mulher, o fator de descapacitação é retirado e ocorre o processo de capacitação, no qual o espermatozoide está apto a fertilizar o ovócito e transferir sua carga de DNA.
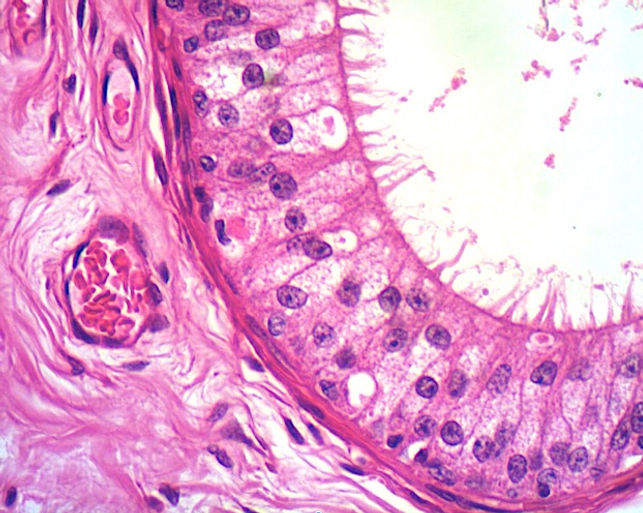
O epitélio do ducto do epidídimo é caracterizado por ser um epitélio pseudoestratificado cilíndrico alto e contendo estereocílios e circundado por uma camada de músculo liso. De modo geral, é composta por dois tipos de células as células principais e as células basais.
-
Células principais: possuem numerosas microvilosidades longas e modificadas, chamadas esterocílios, são estruturas imóveis e que aumentam a superfície de contato. Atuam tanto na reabsorção quanto na secreção. A maior parte do líquido não reabsorvido pelos dúctulos eferentes sofre reabsorção na parte proximal do epidídimo. As células epiteliais também fagocitam quaisquer corpos residuais que não foram removidos pelas células de Sertoli, bem como os espermatozoides que degeneraram no ducto. Devido a essa função absortiva, são encontrados diversos vacúolos e canalículos em seu citoplasma (aparecem esbranquiçados na lâmina). Além disso, secretam fatores, como a glicerofosfocolina, ácido siálico, glicoproteínas e esteroides que ajudam na maturação dos espermatozoides. As células principais são muito altas na parte inicial do epidídimo e diminuem de altura conforme o epitélio progride em direção à extremidade distal ou cauda do epidídimo.
-
Células basais: estão sobre a lâmina basal e possuem formato esférico. Fazem parte das células-tronco do epitélio ductal.
As células epiteliais do ducto epididimário estão apoiadas sobre uma lâmina basal que é recoberta por células musculares lisas e por tecido conjuntivo frouxo. A camada de músculo liso do ducto do epidídimo aumenta gradualmente em espessura, adquirindo três camadas na região da cauda. Na cabeça do epidídimo e em boa parte do corpo, o músculo liso compõe uma fina camada disposta de modo circular, lembrando a estrutura dos ductos eferentes. Em contrapartida, na cauda do epidídimo, são adicionadas duas camadas longitudinais, uma longitudinal interna e outra longitudinal externa. As três camadas tornar-se-ão contínuas com as três camadas de músculo liso presente no ducto deferente, a continuação do sistema de ductos excretores.
Sob esse viés, a adição de camadas de músculo liso está diretamente associada à sua função, por exemplo, na cabeça e no corpo do epidídimo, as contrações peristálticas têm como intuito mover os espermatozoides ao longo do ducto. Já as contrações na cauda do epidídimo, principal reservatório de espermatozoides maduros, está associada a estimulação neural referente ao estímulo da ejaculação.
EPIDÍDIMO - H.E. - 40x
EPIDÍDIMO - H.E. - 100x
EPIDÍDIMO - H.E. - 400x
4. DUCTO DEFERENTE
Cada testículo está envolto por uma camada espessa de tecido conjuntivo denso modelado, a túnica albugínea. Ela é mais densa na superfície posterior dos testículos para formar o mediastino do testículo, do qual se originam os delgados septos fibrosos. Os septos ao penetrarem no testículo, divide-o em compartimentos piramidais chamados lóbulos testiculares. Cada testículo é dividido em aproximadamente 250 lóbulos que se originam a partir dessa cápsula de tecido conjuntivo. Esses septos são incompletos e muitas vezes há comunicação entre os lóbulos. Cada lóbulo do testículo é composto de um a quatro túbulos seminíferos, local onde os espermatozoides são produzidos. Os túbulos seminíferos são envolvidos por um tecido conjuntivo frouxo rico em vasos linfáticos e sanguíneos, nervos e células de Leydig (intersticiais) que são as responsáveis pela produção de testosterona. Além disso, o tecido intersticial do testículo é indispensável para a nutrição das células dos túbulos seminíferos, a produção de andrógenos e o transporte destes hormônios para a circulação sistêmica.